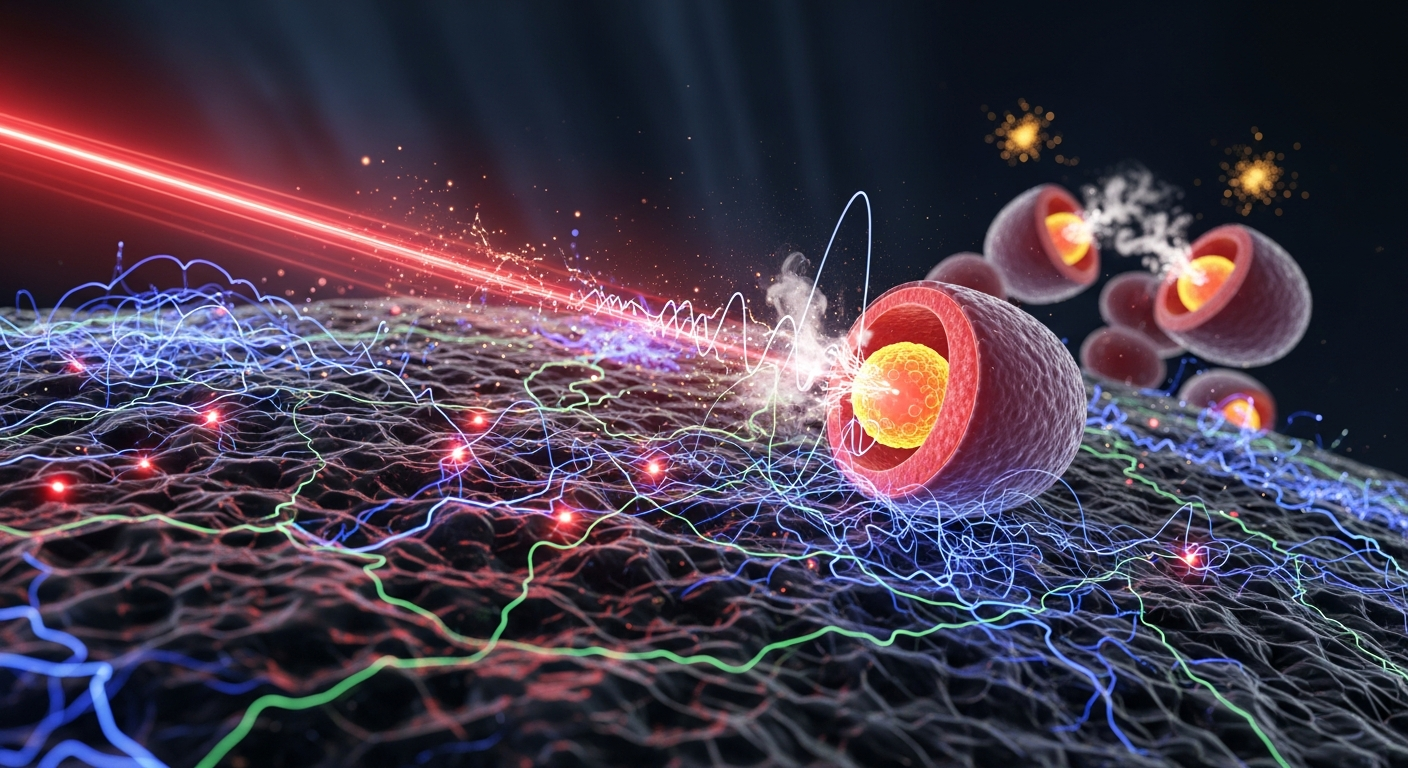
Photobiomodulation therapy, a non-invasive treatment using red and near-infrared light, has demonstrated efficacy in applications from wound healing to neurorehabilitation. Yet, a critical variable remains underexplored: melanin. This article explores how melanin's unique biophysical properties not only influence the delivery of light to target tissues but may also position it as an active participant in the cellular response to photonic energy.
A peculiar and defining characteristic of photobiomodulation (PBM) is its biphasic dose-response curve. Within a specific range of light intensity and duration, cellular functions like proliferation and repair are stimulated. Outside this "therapeutic window," however, the effect diminishes rapidly and can even become inhibitory. This phenomenon, known as the Arndt-Schulz law, suggests a delicate biological balancing act. But what happens when a primary biological variable—the concentration of melanin in the tissue—is introduced? The straightforward calculation of light dosage becomes profoundly more complex. Melanin, a molecule with one of the broadest absorption spectra in nature, acts as a potent optical filter, fundamentally altering the energy that reaches deeper cellular targets. This raises a crucial question for the future of photomedicine: Is melanin merely a passive shield, or is it an active player in transducing light into biological information?
The Canonical Mechanism: Light and the Mitochondrial Powerhouse
To understand melanin’s role, we must first examine the established mechanism of PBM. The prevailing scientific consensus, built on decades of research by pioneers like Tiina Karu, identifies a specific molecule within our cells' mitochondria as the primary photoacceptor for red and near-infrared (NIR) light: cytochrome c oxidase (CCO), also known as Complex IV of the mitochondrial electron transport chain. CCO contains copper and heme chromophores that exhibit absorption peaks in the red and NIR regions of the electromagnetic spectrum.
The mechanism proceeds as follows: under conditions of cellular stress or hypoxia, nitric oxide (NO) can bind to CCO, competitively inhibiting oxygen and slowing down the electron transport chain. This leads to a decrease in the production of adenosine triphosphate (ATP), the cell's primary energy currency, and an increase in reactive oxygen species (ROS), which can cause oxidative damage. When photons of the correct wavelength (typically 630-900 nm) are absorbed by CCO, they are thought to photodissociate the inhibitory nitric oxide. This "re-activates" the enzyme, restoring oxygen consumption, increasing the mitochondrial membrane potential, and boosting ATP synthesis. The brief, controlled burst of ROS that follows also functions as a crucial signaling molecule, activating transcription factors like NF-κB and AP-1. These factors then orchestrate a cascade of downstream genetic expression, leading to increased cell proliferation, reduced inflammation, and enhanced tissue repair. This mitochondrial-centric model provides a robust explanation for many of PBM's observed therapeutic effects.
Melanin: A Broadband Competitor for Photonic Energy
The mitochondrial model is elegant, but it implicitly assumes that photons arrive at their CCO target unimpeded. In biological reality, this is rarely the case, especially in pigmented tissues. Melanin, particularly the dark brown-black polymer eumelanin, is an exceptionally efficient absorber of photons across a vast spectrum, including the ultraviolet, visible, and near-infrared ranges. While its absorption coefficient decreases as wavelength increases, it remains a significant chromophore throughout the PBM therapeutic window.
This presents a direct biophysical challenge: melanin competes with cytochrome c oxidase for the same incident photons. Research by Meinhardt, Holleran, and others has quantified this effect. In individuals with higher concentrations of epidermal melanin (i.e., darker skin phototypes on the Fitzpatrick scale), a significant fraction of the light energy is absorbed in the upper layers of the skin before it can penetrate to dermal or subcutaneous targets where mitochondria in fibroblasts, macrophages, or muscle cells reside. This optical shielding effect means that a PBM dose that is therapeutic for fair skin may be sub-therapeutic for highly pigmented skin, as insufficient photonic energy reaches the target depth. Conversely, attempting to compensate by increasing the light intensity at the surface raises the risk of inducing thermal effects, as the absorbed energy is primarily converted to heat. This dynamic fundamentally complicates dosimetry and is a critical factor in the variable outcomes reported in some PBM clinical trials.
Beyond a Passive Shield: Melanin as a Bioelectric Transducer
The role of melanin as a passive optical filter is well-established, but its unique material properties suggest a far more active and intriguing role. Research at the Quantum Melanin Research Foundation and other institutions is exploring melanin's capabilities as an organic semiconductor and a proton conductor. John McGinness and his colleagues first proposed in the 1970s that melanin could function as an amorphous semiconductor, capable of absorbing photonic energy to excite electrons and generate charge carriers. This property is not merely theoretical; it is leveraged in nascent organic electronics. In a biological context, this raises a compelling hypothesis: Could melanin be transducing absorbed light energy not just into heat, but into electrical or chemical signals?
Consider melanin’s interaction with water. Melanin is highly hydrated, and its electrical conductivity is known to be dependent on its hydration state. It exhibits significant proton conductivity, meaning it can facilitate the movement of protons (H+ ions). This is a foundational process in cellular bioenergetics, most famously in the mitochondrial membrane where proton gradients drive ATP synthesis. Could melanin, upon absorbing light, use that energy to locally alter proton gradients or generate micro-electrical fields?
This theoretical framework connects PBM to the field of bioelectricity, which studies how cells use ion flows and electrical potentials to direct growth, regeneration, and pattern formation. Work from the laboratory of Michael Levin at Tufts University has demonstrated that cellular collectives read and respond to bioelectric cues to build and repair complex anatomical structures. If melanin can convert light into a bioelectric signal, it could potentially influence cellular behavior through mechanisms complementary to the CCO pathway. It might modulate membrane potentials, influence ion channel activity, or interact with the cell’s endogenous electric fields. This view recasts melanin from a simple sunblock into a potential photo-bioelectric interface, a component of a larger cellular information processing network. This remains an area of frontier science, but one that is critical to a complete understanding of light-tissue interactions.
Key Takeaways
- The primary accepted mechanism for photobiomodulation involves the absorption of red and near-infrared light by cytochrome c oxidase in mitochondria, which enhances ATP production and triggers beneficial signaling cascades.
- Melanin is a powerful, broadband chromophore that directly competes with mitochondrial targets for photons, acting as an optical shield that reduces the penetration of light into deeper tissues.
- The concentration of melanin in the skin is a critical variable for PBM dosimetry; higher melanin levels may require adjustments in wavelength or fluence to achieve a therapeutic effect at a target depth without causing surface heating.
- Beyond passive absorption, melanin's inherent properties as a hydrated, amorphous semiconductor and proton conductor suggest it may actively transduce light energy into bioelectric or biochemical signals.
- The potential for melanin to act as a photo-bioelectric interface is a frontier research area that could bridge photomedicine with the science of bioelectric signaling in development and regeneration.
- A comprehensive understanding of PBM requires moving beyond a single-chromophore model to account for the complex interplay between light, mitochondria, and melanin-rich structures.
References
- Karu, T. I. "Mitochondrial signaling in low-level light therapy." Neurophotonics 2(3), 031201 (2015). DOI: 10.1117/1.NPh.2.3.031201.
- Hamblin, M. R. "Mechanisms and applications of the anti-inflammatory effects of photobiomodulation." AIMS Biophysics 4(3), 337–361 (2017). DOI: 10.3934/biophy.2017.3.337.
- Jacques, S. L. "Optical properties of biological tissues: a review." Physics in Medicine & Biology 58(11), R37–R61 (2013). DOI: 10.1088/0031-9155/58/11/R37.
- McGinness, J., Corry, P., & Proctor, P. "Amorphous semiconductor switching in melanins." Science 183(4127), 853–855 (1974). DOI: 10.1126/science.183.4127.853.
- Levin, M., Pezzulo, G., & Fiston-Lavier, J. "The Taming of the Shrew: The Composer’s Guide to the Interpretation of Morphogenetic Music." BioEssays 33(7), 511-523 (2011). DOI: 10.1002/bies.201100030.
- Avci, P., Gupta, A., Sadasivam, M., Vecchio, D., Pam, Z., Pam, N., & Hamblin, M. R. "Low-level laser (light) therapy (LLLT) in skin: stimulating, healing, restoring." Seminars in Cutaneous Medicine and Surgery 32(1), 41–52 (2013).
- Solís Herrera, A., Arias Esparza, M. C., Solís Arias, R. I., Solís Arias, M. P., & Solís Arias, C. "The unexpected capacity of melanin to dissociate the water molecule fills the gap between the life before and after the appearance of photosynthesis." International Journal of Hydrogen Energy 35(21), 12079-12085 (2010). DOI: 10.1016/j.ijhydene.2010.08.067.
- Tafur, J., & Mills, P. J. "Low-intensity light therapy: exploring the role of redox mechanisms." Photomedicine and Laser Surgery 26(4), 323–328 (2008). DOI: 10.1089/pho.2007.2184.